발달 초기의 배아는 둥근 공 모양입니다. 이런 배아가 발달하면서 머리-꼬리(anterior-posterior), 배-등(ventraldorsal), 왼쪽-오른쪽(left-right) 같은 몸의 축이 만들어집니다. 머리-꼬리 축을보면 이 축을 따라 다양한 몸의 기관들이 발달한다는 점을 알 수 있습니다. 척추만 보더라도 가장 위에 목뼈, 아래로 내려가면서 등뼈, 허리뼈, 엉치뼈, 꼬리뼈 차례로 구성돼 있죠. 이렇게 축을 따라서 특정한 구조가 발달하는 것을 ‘신체의 패턴화(body patterning)’라고 부릅니다.
몸의 ‘패턴’을 결정하는 혹스 유전자
우리 몸이 어떻게 패턴화되는지에 대한 연구는 초파리에서 시작합니다. 과일 껍질을 제대로 버리지 않고 놔두면 모여드는 작은 파리들이 바로 초파리인데요. 주방에서는 골칫덩어리일지 모르지만 유전학과 발생학 등의 연구 분야에서는 그만한 효자생물이 없습니다.
과학자들은 정상과 다른 돌연변이 초파리를 관찰하던 중 충격적인 모습의 돌연변이 초파리 한 마리를 발견했습니다. 바로 머리 위 더듬이가 있어야 할 자리에 다리가 있는 돌연변이였죠(아래 사진). 훗날 과학자들은 다리쪽에서 발현되는 ‘안테나페디아(Antennapedia)’ 유전자가 돌연변이로 인해 머리쪽에서 발현되면 머리에서 다리가 만들어진다는 사실을 밝혀 냈습니다.
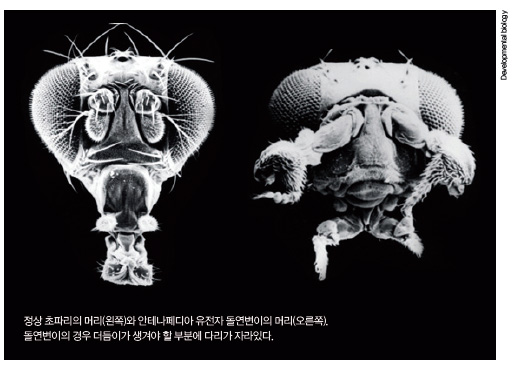
더 놀라운 사실은 안테나페디아 유전자처럼 신체의 패턴화를 담당하는 유전자들이 더 있다는 겁니다. 게다가 이 유전자들은 염색체 여기저기에 분산돼 있는 게 아니라 초파리의 염색체 하나에 옹기종기 모여 있었습니다. 이런 유전자들을 ‘혹스 유전자(hox genes)’라고 부릅니다. 혹스 유전자들의 중요한 특징은 ‘공통 직선성(colinearity)’입니다. 하나의 염색체에 모여 있는 혹스 유전자들을 가장 앞에 있는 유전자부터 가장 끝에 있는 유전자까지 쭉 나열하면, 앞에 있는 유전자부터 순서대로 머리, 목, 가슴, 꼬리 부분의 구조를 만들어내는 기능을 한다는 뜻입니다(아래 그림). 염색체 상의 유전자 위치와 머리-꼬리 축에서 기능을 할 위치가 서로 대응된다는 것이죠.
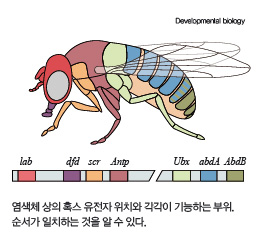
혹스 유전자는 초파리에만 존재하는 것이 아닙니다. 인간을 포함한 포유류의 배아 발달에도 혹스 유전자가 큰 역할을 합니다. 초파리에는 여러 개의 혹스 유전자가 하나의 그룹으로 묶여 있는데 진화 과정 동안 복제가 일어나면서 쥐와 인간의 유전체에는 이런 그룹이 4개나 존재 합니다.
혹스 유전자에 돌연변이가 생긴 쥐는 초파리와 마찬가지로 이상한 신체 패턴을 보이는데요. 예를 들어 ‘HOXD3’ 유전자가 제 기능을 하지 못하면 첫 번째 목뼈가 정상적으로 발달하지 못해 두개골과 부분적으로 붙어 버립니다. 혹스 유전자의 정상적인 발현을 방해하면 팔과 다리의 위치가 바뀔 수도 있고, 팔이 4개가 만들어질 수도 있습니다.

인간 정자의 길이는 약 0.05mm, 현미경 없이는 볼 수 없는 크기입니다. 반면 난자의 지름은 0.1~0.15mm입니다. 정자보다 2~3배가량 크죠. 난자의 세포질 안에는 각종 세포소기관을 비롯해 RNA와 단백질이 가득 차 있습니다. 따라서 수정란의 유전물질은 아빠의 유전물질 반, 엄마의 유전물질 반이지만 수정란의 세포질은 모두 난자의 것이라고 봐도 무방합니다.
수정 뒤 첫 세포분열, 난자 속 RNA 덕분
수정란은 배아로 발달하기 위해 세포분열을 시작합니다. 그런데 이때 수정란 안에있는 DNA는 단백질을 만들어내지 못하는 상태입니다. 그럼에도 수정란이 어떻게 세포분열을 하고 생존할 수 있는지 의아해지죠?
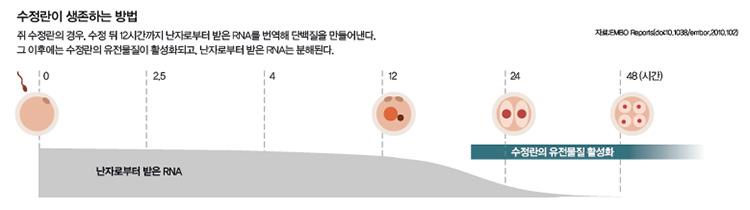
비밀은 난자로부터 받은 세포질에 있습니다. 수정란은 수정 뒤 몇 번의 세포분열을 하는 동안 자신의 유전물질이 아닌, 난자로부터 받은 세포질에 저장된 RNA로 단백질을 만들어 생존합니다. 그 기간은 생물마다 조금씩 다릅니다. 개구리의 경우 12회, 제브라피시는 10회, 쥐의 경우 1회의 세포분열을 하는 동안 난자에게 신세를 집니다.
본래 난자 안에 저장돼 있는 RNA는 일부가 비활성화된 상태입니다. RNA의 꼬리 부분에 특정 단백질이 붙어서 RNA가 단백질로 번역되는 것을 막고 있기 때문입니다. 그런데 수정이 이뤄짐과 동시에 이 단백질들이 RNA에서 떨어져나갑니다. 이렇게 활성화된 RNA가 단백질로 번역돼 수정란의 세포분열을 돕습니다.
또 세포분열이 일정 횟수 이상 일어나면 수정란의 세포질에 저장돼 있던 RNA는 분해되기 시작합니다. 반대로 수정란의 유전물질이 활성화되기 시작하는데요. 어떻게 이렇게 기가 막힌 타이밍을 잡는지는 아직 명확하게 밝혀지지 않았습니다.
유력한 가설 중 하나는 수정란의 DNA활성화를 막는 억제 단백질이 있는데, 이것이 세포분열 과정에서 희석돼 유전자 발현이 시작된다는 가설입니다. 예를 들어 수정란에 억제 단백질 8개가 DNA를 꽁꽁 묶고 있다고 생각해 봅시다. 세포분열 이후 수정란이 8세포기로 접어들면 세포 각각에 있는 억제 단백질은 1개로 급격히 줄어듭니다.
한편 세포분열 횟수와 관계없이 수정란이 수정된 지 몇 분이 지났는지를 확인해 일정 시간이 지나면 자신의 유전물질을 활성화시킨다는 가설도 있습니다.
또 다른 가설로는 발생 초기에는 세포분열이 너무 빨라서 유전물질이 발현될 시간이 부족하고, 세포분열의 속도가 느려지면 비로소 유전물질이 발현된다는 주장도 있습니다.
어쨌든 분명한 것은 수정란의 유전물질이 활발하게 발현되는 시기와 난자의 RNA가 분해되는 시기가 겹친다는 사실입니다. 분해되는 과정은 아직 밝혀지는 중입니다만, 수정란의 유전물질이 만들어낸 짧은 RNA 하나가 난자의 RNA 꼬리에 붙어서 난자의 RNA를 야금야금 분해시키는 효소를 불러온다는 연구도 있습니다(doi:10.1126/science.1122689). 이 일련의 과정이 수정란이 자궁에 착상하기도 전에 일어나는 일이라고 하니 정말 놀랍지 않습니까.