1930년대 러시아의 식물학자 미하일 샤일라얀은 희한한 실험을 했다. 꽃이 핀 식물의 잎을 잘라 아직 꽃피지 않은 식물에 접목하자 이 식물도 꽃이 피었던 것. 이런 현상은 종이 다른 식물 사이에서도 일어났다. 샤일라얀은 잎에서 꽃이 피게 하는 신호물질이 만들어진 뒤 꽃봉오리가 생기는 부분으로 이동해 꽃이 피게 한다고 가정하고 이 가상의 물질을 ‘화성소(florigen)’라고 불렀다. 그 뒤 70여 년 동안 수많은 식물학자들이 화성소의 실체를 밝히기 위해 헌신했고 좌절했다.
그런데 1999년 12월 3일자 과학저널 ‘사이언스’에 개화유전자 ‘FT’의 발견에 대한 논문이 실리면서 화성소의 실체가 드러나기 시작했다. 물론 이 논문이 발표됐을 때만 해도 저자들조차 자신들이 발견한 유전자가 화성소의 유전자임을 알지 못했지만.
고려대 안지훈 교수, 주저자로 참여
개화유전자란 식물의 꽃이 피는 데 관여하는 유전자로 1990년대부터 본격적으로 연구되기 시작했다. 연구자들은 식물 유전체에 DNA를 변형시키는 화학약품을 처리해 인공적인 돌연변이를 일으켰을 때 개화시기가 늦어지거나 빨라지는 변화를 보이는 돌연변이체를 여러 개 찾아냈다. 만일 어떤 돌연변이체가 꽃이 늦게 핀다면 아마도 개화를 촉진하는 유전자가 고장 난 결과라고 추정할 수 있다.
개화 돌연변이체들의 염색체에서 변이가 일어난 위치를 추적한 결과 개화 과정에 여러 유전자가 관여한다는 사실이 밝혀졌다. 이처럼 다양한 유전자들이 개화조절에 참여하는 이유는 변화하는 외부 환경 조건이나 성장상태를 반영해 최적의 타이밍에서 꽃을 피우기 위해서다. 식물이 생식기관인 꽃을 언제 피우느냐는 종이 살아남는 데 결정적인 역할을 한다.

미국 솔크연구소 식물생물학실험실의 데트레프 바이겔 박사(현재 독일 튀빙겐 막스플랑크연구소 책임자)팀은 개화유전자를 찾기 위해 ‘활성화표지법(activation tagging)’이란 새로운 방법을 도입했다. 활성화표지법이란 숙주의 유전체에 끼어들어가 주변의 유전자 전사를 활성화하는 바이러스의 DNA조각(‘전사 인핸서(enhancer)’라고 부른다)을 이용하는 방법이다. 즉 전사 인핸서를 식물세포 안에 넣으면 인핸서가 식물유전체 안으로 들어간다. 이 식물의 씨를 얻어 뿌리면 전사 인핸서가 활성화시킨 유전자에 따라 다양한 특징을 보이는 식물체가 만들어진다.
예를 들어 개화유전자 부근에 전사 인핸서가 들어갔다면 개화유전자가 활성화돼 그 결과 식물체는 꽃이 빨리 핀다. 이런 식물체를 선별한 뒤 집어넣은 DNA 조각이 유전체의 어느 부분에 들어갔는지 추적해(인핸서의 염기서열을 알고 있으므로 쉽게 찾을 수 있다) 활성화된 주변 유전자를 밝히는 방법이다. 반면 화학약품으로 돌연변이를 일으키는 기존의 방법은 염색체에서 변이가 일어난 대략적인 부분은 쉽게 알 수 있지만 고장 난 유전자를 정확히 찾기가 매우 어렵다.
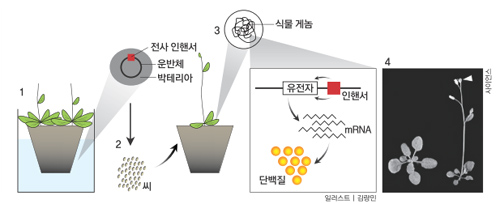
이 연구에는 당시 솔크연구소에서 박사후연구원으로 있던 고려대 생명과학부 안지훈 교수가 참여했다. 안 교수는 논문의 제1저자 3명 가운데 한명이다. 연구자들이 실험에 사용한 식물은 십자화과 1년생 풀인 애기장대다. 애기장대는 싹이 튼 뒤 자라면서 새로운 잎이 계속 나오는데, 보통 14장쯤 되면 꽃대가 올라오고 자그마한 흰 꽃이 핀다. 그런데 활성화표지법으로 얻은 ‘1733’이라고 명명한 식물체는 잎이 4장쯤 됐을때 벌써 꽃이 펴 버렸다. 연구자들은 이 개체에서 전사 인핸서 위치를 알아낸 뒤 주변 유전자의 전사 정도를 확인했다.
그 결과 전사 인핸서 옆에 있는 조그만 단백질 정보를 담고 있는 유전자의 전령RNA(mRNA)가 많이 존재한다는 사실을 발견했다. 이 근처가 고장 난 돌연변이체 3종이 이미 알려져 있었는데, 흥미롭게도 모두 꽃이 늦게 피는 특성을 보였다. 이를 발견한 연구자들은 3종에서 공통적으로 고장 났을 것으로 추정되는 유전자를 ‘FT’라고 명명하고 돌연변이체를 소문자(유전자가 고장 났다는 의미)를 써서 각각 ft-1, ft-2, ft-3라고 불렀다. 그러나 당시에는 FT 유전자의 실체를 몰랐다.
바이겔 박사팀은 이들 돌연변이체(꽃이 늦게 핌)에서 고장 난 유전자가 활성화표지법으로 얻은, 1773(꽃이 일찍 핌)의 전사가 증폭된 유전자와 동일할 것이라고 예상하고 후속연구를 진행했다. 그 결과 실제로 세 돌연변이체 모두 이 유전자가 손상돼 있음을 확인했다. 또한 ft 돌연변이체에 이 유전자를 넣어주자 늦게 꽃피는 특성이 사라졌다. 돌연변이체에서 손상된 유전자와 1733에서 활성화된 유전자가 동일하다는 결정적인 증거였다.
연구결과 FT는 개화과정 네트워크에서 있어서 핵심적인 역할을 한다는 사실이 밝혀졌다. 또 현화식물(속씨식물), 즉 꽃피는 식물은 모두 애기장대의 FT에 해당하는 유전자가 있다는 사실도 밝혀졌다. 그러나 이끼나
고사리처럼 꽃이 피지 않는 식물에는 FT에 해당하는 유전자가 없다. 또 하나 흥미로운 사실은 꽃이 피기 직전 FT 유전자가 활성화되는 부분이 바로 잎이라는 점이다. 이는 샤일라얀이 제안한 화성소의 조건 하나를 만족시키는 특성이다.
거짓으로 밝혀진 2005년 10대 뉴스
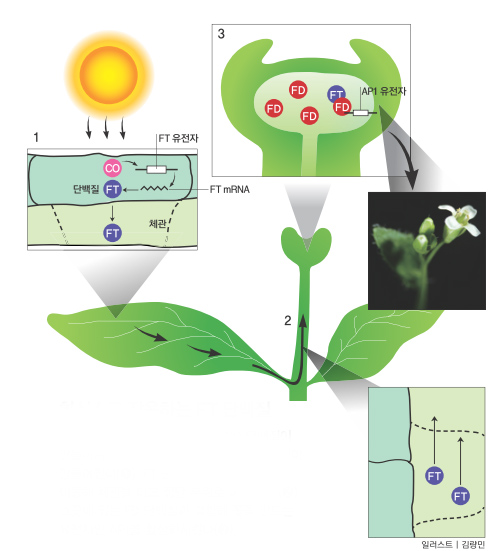
2005년 9월 9일자 ‘사이언스’에는 70년간 찾아 헤맸던 화성소의 실체가 마침내 밝혀졌다는 깜짝 놀랄 만한 연구결과가 실렸다. 그 실체는 바로 FT의 mRNA. 스웨덴농업과학대 우베 닐슨 교수팀은 논문에서 잎에서 만들어진 FT mRNA가 체관을 통해 꽃봉오리가 생기는 정단 부위로 이동한다는 증거를 제시했다.
놀랍게도 FT의 mRNA는 세포를 빠져 나와 먼 거리를 이동했던 것. FT가 보이는 모든 행동은 먼 거리에서 꽃을 피게 만드는 화성소의 행동과 일치했다. 이들의 연구결과는 각종 매체에서 대서특필됐고 ‘사이언스’가 뽑은 2005년 10대 뉴스에도 들어갔다. 박사후연구원 시절, 이 논문에 제1저자로 참여했던 타오 후앙 박사는 그 뒤 중국 샤먼대에 자리를 잡았다.
그러나 탄성이 실망으로 바뀌는 데는 오랜 시간이 걸리지 않았다. 이스라엘 테크니온이스라엘공대 생물학과 엘리저 리프쉬츠 교수팀은 2006년 4월 18일자 ‘미국립과학원회보(PNAS)’에 발표한 논문에서 토마토에서 FT에 해당하는 유전자인 SFT의 mRNA가 이동하는 현상을 발견하지 못했다고 보고했다. 그 뒤 닐슨 교수는 후앙 박사의 데이터를 면밀히 분석했고 결과가 조작된 흔적을 발견했다.
결국 닐슨 교수팀은 2007년 4월 20일자 ‘사이언스’에 2005년 논문을 철회한다는 편지를 실었다. 이 글에서 연구자들은 “실험을 재현한 결과 FT mRNA가 이동한다는 증거를 찾지 못했다”며 “하지만 제1저자가 논문철회에 강하게 반대해서 철회 요청자 명단에서 뺐다”고 설명했다. 실제로 후앙 박사는 여전히 데이터 조작을 부정하고 있다.

mRNA가 아니라 단백질이 이동
화성소의 실체가 다시 미궁에 빠질 즈음 새로운 연구결과 두 편이 2007년 5월 18일자 ‘사이언스’에 나란히 실렸다. 독일 콜론 막스플랑크연구소의 조지 쿠플랜드 박사팀은 애기장대에서 FT mRNA가 아닌 FT 단백질이 체관을 따라 이동한다는 사실을 밝혀냈다. 함께 게재된 논문에서 일본 나라과학기술연구원 시마모토 고 박사팀은 벼에서 FT에 해당하는 Hd3a의 단백질이 이동한다는 증거를 보고했다. 그 뒤 다른 학술지에도 여러 식물의 FT 해당 유전자의 단백질이 이동한다는 사실이 속속 보고됐다. 화성소의 실체가 마침내 밝혀진 셈이다.
쿠플랜드 박사는 “체관의 액 속에는 FT 단백질 말고도 여러 단백질이 존재한다”며 “이는 식물에서 단백질의 장거리 이동이 신호전달의 수단임을 시사한다”고 설명했다. 1999년 논문에서 FT 유전자의 실체를 밝힌 바이겔 박사는 이들의 연구결과에 대해 “이제 FT 단백질이 화성소의 주된 요소라는 데는 의심의 여지가 거의 없다”고 평가하면서 “하지만 다른 분자들도 이 과정에 참여할 것으로 보인다”고 덧붙였다. 실제로 애기장대에서는 FT 단백질과 매우 비슷한 TSF 단백질이 마찬가지 방식으로 이동해 개화를 촉진한다는 사실이 밝혀졌다.
그런데 잎이 꽃을 피우는 신호를 전달한다면 개나리, 진달래, 벚꽃은 어떻게 잎 없이도 이른 봄에 꽃을 피울 수 있을까. 이런 식물들은 잎이 달려 있는 여름이나 가을에 꽃봉오리가 형성되지만 바로 꽃이 피지 않고 잠복기에 들어간다. 그리고 겨울을 난 뒤 기온이 올라가는 주변 신호를 받아 꽃봉오리가 깨어나면서 꽃이 핀다. 이들도 화성소 이론의 예외가 아닌 셈이다.
아직도 끝나지 않은 논란
FT 단백질이 잎에서 만들어진 뒤 정단 부위로 이동하는 설명은 현재 정설이다. 그런데 FT mRNA가 이동한다는 2005년 논문이 철회되면서 이에 대한 논란이 꺼져가던 2009년 ‘바이러스학저널’ 2월 4일자에 약간 생뚱맞은(?) 논문이 게재됐다.
영국의 워릭대 홍 이궈 박사팀은 식물 바이러스 시스템을 이용해서 연구한 결과 FT mRNA도 여전히 이동한다는 연구 결과를 보고했다. 또한 FT mRNA의 1~102번째 염기가 FT mRNA의 이동에 중요하다는 결과도 보고했다. 이 연구팀은 FT 단백질과 함께 FT mRNA도 식물에서 꽃을 피게 하는 신호의 전파에 관여할 것이라고 주장했다. 과연 이 연구팀이 제시한 결과처럼 FT 단백질뿐 아니라 FT mRNA도 이동할까. 꺼져가는 불씨를 되살리며 새로운 불을 지피는 연구결과가 될지, 헛발질을 한 결과가 될지는 좀 더 지켜봐야 할 것 같다.
현재까지 보고된 연구결과를 종합해보면 FT 단백질의 이동은 부정할 수 없는 사실로 보인다. 그러나 아직도 해결해야 할 의문이 많이 남아 있다. FT 단백질이 식물의 정단 부위까지 이동하는 것은 분명한데, 이 FT 단백질이 실제 꽃을 피게 만드는 결정적인 원인인지에 대한 답은 아직 없다. 또 이동한 FT 단백질이 어떻게 꽃봉오리까지 개화 신호를 전달하는지, 체관 내에서 어떻게 이동하는지, 이동하는 과정에서 개화신호를 선별하는 과정이 있는지 궁금한 점이 매우 많다. 야심 찬 후속세대의 과학에 대한 열정만이 이 의문을 풀 것이다.
글 안지훈 고려대 생명과학부 교수 jahn@korea.ac.kr