정보를 저장하기 위해서는 언어가 필요하다. 컴퓨터가 사용하는 기계어는 0과 1로 이뤄진 이진법 숫자다. 세포는 A, G, T, C 등 네 가지 염기로 DNA에 기록을 남긴다. 후성유전도 세포에 DNA 사용법을 알려주고 기록하기 위한 언어가 필요하다. 후성유전의 언어는 DNA 메틸화와 히스톤 변형이다.
0DNA 메틸화는 후성유전학보다 더 유명하다. 후성유전학의 정확한 의미를 모르는 사람도 DNA 메틸화는 안다. 메커니즘이 그만큼 간단하기 때문이다. 염색체나 히스톤 같은 복잡한 개념을 몰라도 ‘DNA에 뚜껑을 씌워 전사를 막는다’는 설명은 꽤 쉽고 그럴듯하게 들린다. 보다 정확하게 설명하면, 메틸화는 DNA 염기 시토신(C)의 육각형 탄소 고리 중 5번 탄소의 수소를 메틸기(-CH3)로 바꾸는 작용이다. 메틸기는 탄소 하나와 수소 세 개로 이뤄진 간단한 형태의 탄소 작용기다. 시토신이 메틸화되면 전사에 참여하는 요소들이 그 자리에 결합하기 어려워져 전사효율이 낮아진다. 하지만 이렇게 간단하고 직관적인 정의와 달리 실제 생명현상에서 메틸화의 역할은 훨씬 복잡하다.
전사를 조절하는 DNA의 섬
후성유전학자들은 인간 유전체 내의 메틸화 분포와 그 역할에 대해 관심을 갖고 있다. 특히 ‘CpG 섬(CpG island)’이라는 게 있다. CpG란 DNA 염기 중 시토신 뒤에 구아닌(G)이 나오는 특별한 반복 서열을 말한다. 이 서열은 인간을 비롯한 포유류에서 특별히 메틸화가 잘 일어나는 부위다. 인간 유전체 전체의 시토신 메틸화 비율은 3% 정도인데, CpG 시토신의 메틸화 비율은 60~90%에 이른다.
CpG 서열은 인간 유전체에서 섬이라고 불릴 정도로 군집을 이루고 있는 경우가 많다. 작게는 200개에서 길게는 3000개에 달하는 구간에서 CpG가 평범한 서열에 비해 50배 이상 자주 반복된다. 흥미로운 점은 전사를 촉진하는 서열인 프로모터에서 CpG 섬이 자주 발견된다는 것이다. 인간 유전자의 프로모터 중 약 70%가 CpG 섬을 포함하고 있다. 평상시 프로모터의 CpG 섬 메틸화 비율은 다른 CpG 서열 보다 낮은 편이다. 전사인자 등이 결합해 전사를 촉진해야 하는 프로모터 특성상 메틸기가 전사를 방해할 수 있기 때문이다.
만약 CpG 섬에 메틸화가 일어나면 프로모터가 비활성화 돼 전사가 일어나지 않는다(침묵). 유전체 각인, X염색체비활성화(3파트 참조) 등 세포는 메틸화를 이용해 의도적으로 유전자를 침묵시키기도 하지만, 질병이 이를 악용하기도 한다. 예를 들어 암은 유전자 전체 수준에서 메틸화를 감소시키지만 CpG 섬의 메틸화 비율은 높여 정상 유전자의 전사를 막는다. 현재 후성유전학자들은 이런 비정상적인 세포의 활동과 CpG 섬의 관계, CpG 섬의 메틸화에 따른 전사요소의 반응 등을 연구하고 있다.

여섯 번째 염기를 찾아서
메틸화는 현재까지 발견된 후성유전 언어 중 가장 지속시간이 길다. 세포에서 다음 세대의 세포로 전달되는 게 확실하고, 생식세포를 통해 후대에도 전달될 수 있다. 때문에 메틸화된 시토신(5mC)을 A,G,T,C에 이은 다섯 번째 염기로 추가해야 된다는 주장이 나오기도 한다. 최근에는 여기에 여섯 번째 염기 후보까지 등장했다. 미국 록펠러대 연구팀은 5mC의 메틸기 수소 하나가 하이드록시기(-OH)로 치환된 하이드록시메틸 시토신(5hmC)을 인간과 쥐의 뇌세포에서 발견해 2009년 ‘사이언스’에 발표했다. 50년 전에 박테리오파지에서 처음 발견됐지만 인간에서 발견된 것은 2009년이 처음이다. 2년 뒤에는 미국 하버드대 연구팀이 TET 단백질이 5mC를 산화시켜 5hmC를 만든다는 것을 밝혀냈다.
아직까지 5hmC가 정확히 어떤 기능을 하는지는 알려져 있지 않다. 특별한 기능 없이 단순한 경유지에 불과하다는 주장도 있다. 5mC가 5hmC로 변하고, 5hmC가 다시 시토신으로 돌아오는 메커니즘이 2011년 발견됐기 때문이다. 하지만 이에 반대되는 증거도 만만치 않다. 일례로 5hmC는 줄기세포나 증폭자 등 중요한 부위에서 자주 발견된다. 올해 6월에는 영국 케임브리지대 연구팀이 5hmC의 변형물 중 하나인 5fC가 존재 비율은 낮지만 발생에 따라 항상 일정한 패턴을 유지한다는 사실을 발견하기도 했다. 만약 이들의 진짜 기능이 발견된다면 후성유전학 언어는 새로운 격변기를 맞게 될 것이다.
유전체의 책갈피, 히스톤 변형
메틸화는 세포를 넘어서 전달될 정도로 강력한 표식이지만 그 범위가 좁다. 유전정보가 아주 두꺼운 책이라면 형광펜으로 한두 단어를 표시하는 정도다. 일분일초가 중요한 초기 발달이나 면역 반응 같은 경우는 메틸화만으로 광범위한 유전체를 빠르게 조절하기 힘들다. 보다 넓은 범위에서 DNA 사용법을 결정하는, 마치 두꺼운 책의 책갈피 역할을 하는 표식이 필요하다. 새로운 표식을 찾기 위해서는 염색체의 복잡한 구조부터 이해해야 한다.
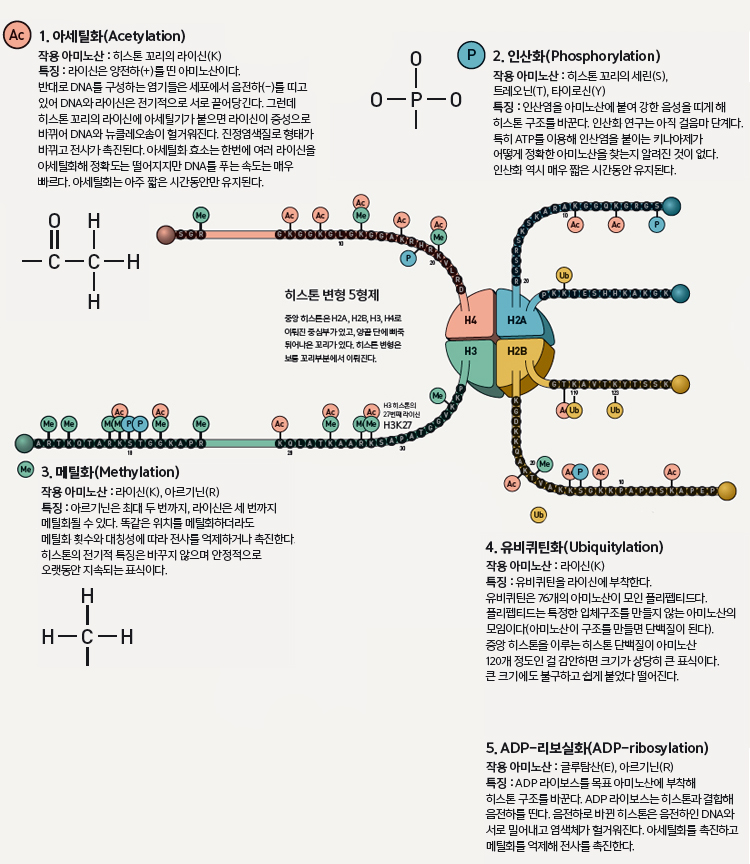
30억 쌍이 넘는 인간 DNA 염기 서열을 일렬로 쭉 펴면 길이만 1.8m다. 문제는 이렇게 긴 DNA를 0.000001m 내외의 핵 안에 넣어야 된다는 것이다. 이를 위해 세포는 실패에 실을 감듯 DNA를 히스톤이라는 단백질에 묶어 보관한다(이 구조를 뉴클레오솜이라고 한다). 이때 실패 역할을 하는 동그란 공처럼 생긴 ‘중앙 히스톤’은 H2A, H2B, H3, H4 히스톤이 두 개씩 결합한 팔량체로, 150여 개의 DNA가 이 팔량체에 감긴다. 이외에 H1 히스톤은 DNA가 들어오고 나가는 곳에서 실패를 고정시키는 집게 역할을 하고, 80여 개의 연결 DNA는 뉴클레오솜과 뉴클레오솜의 사이를 잇는다. 이런 뉴클레오솜이 촘촘히 뭉쳐 염색체가 된다. 전사를 시작하기 위해서는 전사인자나 증폭자가 프로모터 지역에 결합해 전사효소를 불러와야 한다. 뉴클레오솜이 오밀조밀 뭉쳐있다면 전사인자가 목표 서열에 결합할 수 없다. 이렇게 전사가 어려운 빡빡한 염색체를 이질염색질(heterochromatin)이라고 부른다. 반대로 뉴클레오솜 사이에 간격이 충분해 전사가 용이한 상태는 진정염색질(euchromatin)이라고 한다.
유전체 수준에서 전사를 조절하려면 필요에 따라 두 가지 상태를 조절할 수 있어야 한다. 특히 중요한 것은 히스톤의 화학적 변형이다. 예를 들어 H3 히스톤의 27번째 라이신(K)에 메틸기가 세 개 붙으면 이질염색질로 변한다(상단 그림 참조). 이외에도 히스톤 아미노산에 아세틸기, 유비퀴틴, 인산 등이 붙고 떨어짐에 따라 뉴클레오솜은 물론이고 염색체 전체의 구조가 바뀌기도 한다. 그중 다섯 가지를 엄선해 소개한다. 이것만 알면 당신도 이제 후성유전 언어의 전문가가 될 수 있다.
▼관련기사를 계속 보시려면?
INTRO. 쌍둥이는 왜 다를까
PART1. 후성유전학이 태어나다
PART2. 세포의 DNA 사용 설명서
PART3. 쓰레기 RNA가 암 정복한다
PART4. 그들이 ‘변신’하는 이유
PART5. 어떻게 살 것인가
0DNA 메틸화는 후성유전학보다 더 유명하다. 후성유전학의 정확한 의미를 모르는 사람도 DNA 메틸화는 안다. 메커니즘이 그만큼 간단하기 때문이다. 염색체나 히스톤 같은 복잡한 개념을 몰라도 ‘DNA에 뚜껑을 씌워 전사를 막는다’는 설명은 꽤 쉽고 그럴듯하게 들린다. 보다 정확하게 설명하면, 메틸화는 DNA 염기 시토신(C)의 육각형 탄소 고리 중 5번 탄소의 수소를 메틸기(-CH3)로 바꾸는 작용이다. 메틸기는 탄소 하나와 수소 세 개로 이뤄진 간단한 형태의 탄소 작용기다. 시토신이 메틸화되면 전사에 참여하는 요소들이 그 자리에 결합하기 어려워져 전사효율이 낮아진다. 하지만 이렇게 간단하고 직관적인 정의와 달리 실제 생명현상에서 메틸화의 역할은 훨씬 복잡하다.
전사를 조절하는 DNA의 섬
후성유전학자들은 인간 유전체 내의 메틸화 분포와 그 역할에 대해 관심을 갖고 있다. 특히 ‘CpG 섬(CpG island)’이라는 게 있다. CpG란 DNA 염기 중 시토신 뒤에 구아닌(G)이 나오는 특별한 반복 서열을 말한다. 이 서열은 인간을 비롯한 포유류에서 특별히 메틸화가 잘 일어나는 부위다. 인간 유전체 전체의 시토신 메틸화 비율은 3% 정도인데, CpG 시토신의 메틸화 비율은 60~90%에 이른다.
CpG 서열은 인간 유전체에서 섬이라고 불릴 정도로 군집을 이루고 있는 경우가 많다. 작게는 200개에서 길게는 3000개에 달하는 구간에서 CpG가 평범한 서열에 비해 50배 이상 자주 반복된다. 흥미로운 점은 전사를 촉진하는 서열인 프로모터에서 CpG 섬이 자주 발견된다는 것이다. 인간 유전자의 프로모터 중 약 70%가 CpG 섬을 포함하고 있다. 평상시 프로모터의 CpG 섬 메틸화 비율은 다른 CpG 서열 보다 낮은 편이다. 전사인자 등이 결합해 전사를 촉진해야 하는 프로모터 특성상 메틸기가 전사를 방해할 수 있기 때문이다.
만약 CpG 섬에 메틸화가 일어나면 프로모터가 비활성화 돼 전사가 일어나지 않는다(침묵). 유전체 각인, X염색체비활성화(3파트 참조) 등 세포는 메틸화를 이용해 의도적으로 유전자를 침묵시키기도 하지만, 질병이 이를 악용하기도 한다. 예를 들어 암은 유전자 전체 수준에서 메틸화를 감소시키지만 CpG 섬의 메틸화 비율은 높여 정상 유전자의 전사를 막는다. 현재 후성유전학자들은 이런 비정상적인 세포의 활동과 CpG 섬의 관계, CpG 섬의 메틸화에 따른 전사요소의 반응 등을 연구하고 있다.
여섯 번째 염기를 찾아서
메틸화는 현재까지 발견된 후성유전 언어 중 가장 지속시간이 길다. 세포에서 다음 세대의 세포로 전달되는 게 확실하고, 생식세포를 통해 후대에도 전달될 수 있다. 때문에 메틸화된 시토신(5mC)을 A,G,T,C에 이은 다섯 번째 염기로 추가해야 된다는 주장이 나오기도 한다. 최근에는 여기에 여섯 번째 염기 후보까지 등장했다. 미국 록펠러대 연구팀은 5mC의 메틸기 수소 하나가 하이드록시기(-OH)로 치환된 하이드록시메틸 시토신(5hmC)을 인간과 쥐의 뇌세포에서 발견해 2009년 ‘사이언스’에 발표했다. 50년 전에 박테리오파지에서 처음 발견됐지만 인간에서 발견된 것은 2009년이 처음이다. 2년 뒤에는 미국 하버드대 연구팀이 TET 단백질이 5mC를 산화시켜 5hmC를 만든다는 것을 밝혀냈다.
아직까지 5hmC가 정확히 어떤 기능을 하는지는 알려져 있지 않다. 특별한 기능 없이 단순한 경유지에 불과하다는 주장도 있다. 5mC가 5hmC로 변하고, 5hmC가 다시 시토신으로 돌아오는 메커니즘이 2011년 발견됐기 때문이다. 하지만 이에 반대되는 증거도 만만치 않다. 일례로 5hmC는 줄기세포나 증폭자 등 중요한 부위에서 자주 발견된다. 올해 6월에는 영국 케임브리지대 연구팀이 5hmC의 변형물 중 하나인 5fC가 존재 비율은 낮지만 발생에 따라 항상 일정한 패턴을 유지한다는 사실을 발견하기도 했다. 만약 이들의 진짜 기능이 발견된다면 후성유전학 언어는 새로운 격변기를 맞게 될 것이다.
유전체의 책갈피, 히스톤 변형
메틸화는 세포를 넘어서 전달될 정도로 강력한 표식이지만 그 범위가 좁다. 유전정보가 아주 두꺼운 책이라면 형광펜으로 한두 단어를 표시하는 정도다. 일분일초가 중요한 초기 발달이나 면역 반응 같은 경우는 메틸화만으로 광범위한 유전체를 빠르게 조절하기 힘들다. 보다 넓은 범위에서 DNA 사용법을 결정하는, 마치 두꺼운 책의 책갈피 역할을 하는 표식이 필요하다. 새로운 표식을 찾기 위해서는 염색체의 복잡한 구조부터 이해해야 한다.
30억 쌍이 넘는 인간 DNA 염기 서열을 일렬로 쭉 펴면 길이만 1.8m다. 문제는 이렇게 긴 DNA를 0.000001m 내외의 핵 안에 넣어야 된다는 것이다. 이를 위해 세포는 실패에 실을 감듯 DNA를 히스톤이라는 단백질에 묶어 보관한다(이 구조를 뉴클레오솜이라고 한다). 이때 실패 역할을 하는 동그란 공처럼 생긴 ‘중앙 히스톤’은 H2A, H2B, H3, H4 히스톤이 두 개씩 결합한 팔량체로, 150여 개의 DNA가 이 팔량체에 감긴다. 이외에 H1 히스톤은 DNA가 들어오고 나가는 곳에서 실패를 고정시키는 집게 역할을 하고, 80여 개의 연결 DNA는 뉴클레오솜과 뉴클레오솜의 사이를 잇는다. 이런 뉴클레오솜이 촘촘히 뭉쳐 염색체가 된다. 전사를 시작하기 위해서는 전사인자나 증폭자가 프로모터 지역에 결합해 전사효소를 불러와야 한다. 뉴클레오솜이 오밀조밀 뭉쳐있다면 전사인자가 목표 서열에 결합할 수 없다. 이렇게 전사가 어려운 빡빡한 염색체를 이질염색질(heterochromatin)이라고 부른다. 반대로 뉴클레오솜 사이에 간격이 충분해 전사가 용이한 상태는 진정염색질(euchromatin)이라고 한다.
유전체 수준에서 전사를 조절하려면 필요에 따라 두 가지 상태를 조절할 수 있어야 한다. 특히 중요한 것은 히스톤의 화학적 변형이다. 예를 들어 H3 히스톤의 27번째 라이신(K)에 메틸기가 세 개 붙으면 이질염색질로 변한다(상단 그림 참조). 이외에도 히스톤 아미노산에 아세틸기, 유비퀴틴, 인산 등이 붙고 떨어짐에 따라 뉴클레오솜은 물론이고 염색체 전체의 구조가 바뀌기도 한다. 그중 다섯 가지를 엄선해 소개한다. 이것만 알면 당신도 이제 후성유전 언어의 전문가가 될 수 있다.
▼관련기사를 계속 보시려면?
INTRO. 쌍둥이는 왜 다를까
PART1. 후성유전학이 태어나다
PART2. 세포의 DNA 사용 설명서
PART3. 쓰레기 RNA가 암 정복한다
PART4. 그들이 ‘변신’하는 이유
PART5. 어떻게 살 것인가