지난 화에서 우리는, 자연 선택이 각 개체의 포괄 적합도를 최대화한다는 사실을 알았다. 즉, 사회적 행동을 하는 개체는 자신의 자식 수(직접 적합도)뿐만 아니라, ‘그 행동을 만드는 유전자를 공유할 확률이 있는 수용자의 자식 수에 끼친 영향의 일부분(간접 적합도)’도 신경을 쓰게끔 행동한다. 그런데 치사하게도(?) 필자는 왜 그렇게 되는지는 알려주지 않았다. ‘협력의 공식’의 핵심을 직관적인 그림 모델로 이해해 보자.
122쪽의 그림을 보며 이야기를 시작해보자. 먼저 사회적 행동이 아닌 형질을 만드는 유전자다(그림1). 예를 들어, 잘 익은 과일처럼 탄수화물이 많은 음식을 달다고 느끼게 하는 유전자는 개체의 생존과 번식에 도움이 된다. 이처럼 개체의 번식 성공도(고전적인 의미의 적합도)를 증가시키는 유전자는 자연 선택돼 빈도가 점차 늘어날 것이다.
반면에 잘 익은 과일을 쓰다고 느끼게 해서 개체의 번식 성공도를 감소시키는 유전자는 도태할 것이다. 사회적 행동이 아닌 형질을 만드는 유전자의 경우, 우리는 그 유전자가 개체에 주는 영향이 (+)인지 (-)인지만 보면 그 유전자가 선택될지를 알 수 있다. 즉, 개체의 적합도는 오직 그 개체의 몸 속에 들어 있는 유전자에 전적으로 달려 있다.
사회적 행동을 만드는 유전자는 골칫거리를 안긴다(그림 2). 번식상의 손실을 감수하면서 무조건 상대에게 번식상의 이득을 주는 이타주의자, 그리고 이득을 받기만 할 뿐 남들에게 이득을 되돌려주지 않는 배신자가 있다고 하자. 이들이 둘씩 짝지어서 상호작용한다고 가정한다. 이때, 각 개체의 적합도는 함께 한 수용자가 누구인가에 따라 달라짐을 알 수 있다.
사회적 행동을 만드는 유전자의 경우, 그 유전자가 개체에 (+) 혹은 (-) 영향을 주는지 알더라도 그 유전자가 선택될지를 알 수 없다. 개체의 적합도는 그 개체 자신뿐만 아니라 주변의 이웃에게도 달려 있기 때문이다. 가수 설현이 평생 자식을 셋 낳았다고 하자. 이 중 일부는 설현의 유전자 덕분이 아니라 설현을 도와준 이웃의 유전자 덕분이다. 마찬가지로, 설현의 이웃이 낳은 자식 가운데 일부는 이웃의 유전자 덕분이 아니라 그 이웃을 도운 설현의 유전자 덕분이다.
이처럼 사회적 행동의 진화에서는 개체가 자신이 평생 낳는 자식수를 온전히 유전적으로 통제하지 못한다. 그러니 개체가 자신의 적합도를 증가시키는 방향으로 선택된다고 결코 말할 수 없다. 이제 해밀턴이 이 문제로 얼마나 속을 썩였을지 짐작이 되는가.
사회적 행동을 설명할 수 있을까
해밀턴은 이 난제를 깔끔하게 해결했다. 갑돌이 몸속에 들어 앉아서 을돌이에게 사회적 행동을 하게 하는 유전자의 관점에서 보면, 갑돌이 혹은 을돌이에게 이득이나 손실을 주는 것과 상관 없이, 그저 자신의 복제본을 다음 세대에 더 많이 남기면 그만이다. 사회적 행동을 만드는 유전자가 남기는 복제본의 개수를 정확하게 세는 방법은 무엇일까. 그림 Ⓒ와 Ⓓ가 보여주듯이, 행위자가 평생 낳는 자식 수만 따지는 (즉, 행위자가 받는 화살표들만 세는) 고전적인 적합도는 탈락이다. 수용자가 누군지에 따라 적합도가 널뛰듯 달라지기 때문이다.
사회적 행동의 탄생
그림 1. 사회적 행동이 아닌 형질의 진화
화살표는 비사회적인 형질을 만드는 유전자가 개체 당사자의 적합도에 영향을 끼침을 뜻한다. 모든 개체는 한 명의 자식을 다음 세대에 남기면 본전이다. 즉, 기본적으로 누구나 부여 받는 적합도는 1이다(꼭 1일 필요는 없다. 편의상 정한 것이며 0 또는 양수면 된다).유전자를 지닌 개체의 적합도가 1보다 크다면 이 유전자는 선택될 것이다. 1보다 작다면 점차 제거될 것이다. Ⓐ유전자가 개체의 적합도를 b만큼 증가시키는 경우. 1+b >; 1이므로 선택된다. Ⓑ유전자가 개체의 적합도를 c만큼 감소시키는 경우. 1-c <; 1이므로 제거된다(b>;0, c>;0).
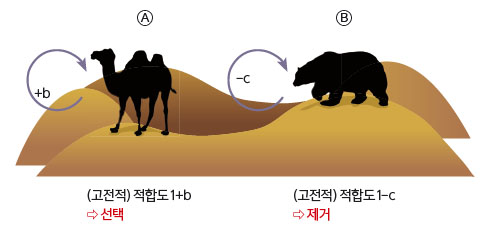
이타주의자는 번식상의 손실(c)을 감수하면서 수용자에게 이득(b)을 준다. 이 때 b>;c>;0, 즉, 두 이타주의자로 구성된 집단이 두 배신자로 구성된 집단보다 전체적으로 더 많은 자식들을 낳는다고 가정한다.
Ⓒ한 이타주의자와 한 배신자로 구성된 집단에서 이타주의자가 거두는 적합도는 1-c다. 이타적 행동에 의한 두 화살표 가운데 남에게 준 화살표는 신경 쓰지 말고 ‘내가 받는’ 화살표만 신경 쓰면 된다. Ⓓ두 이타주의자로 구성된 집단에서, 이타주의자는 수용자에게 협력을 하면서 -c만큼 적합도가 변한다. 또 수용자의 협력 덕분에 +b만큼 적합도가 변한다. 따라서 두 명의 이타주의자로 구성된 집단에서 이타주의자가 거두는 적합도는 1+b-c가 된다(내가 받는 화살표만 신경 쓰시라!). 즉, 수용자가 누구인가에 따라 이타주의자의 적합도가 다르다.
그림 3. 포괄 적합도와 이웃-조정 적합도
Ⓔ포괄 적합도는 행위자의 시각에서 ‘내가 (나 자신을 포함해) 다른 개체들에게 준’ 적합도상의 효과를 각각 유전적 근연도를 가중해서 더한 값이다. Ⓕ이웃-조정 적합도는 수용자의 시각에서 (나 자신을 포함해서) 다른 개체들이 ‘나에게 준’ 적합도상의 효과를 각각 유전적 근연도를 가중해서 더한 값이다. 포괄 적합도와 이웃-조정적합도 모두 1+ 1·(-c) + r·(b)이다. 이것이 1보다 크다면 이 행동은 선택된다.
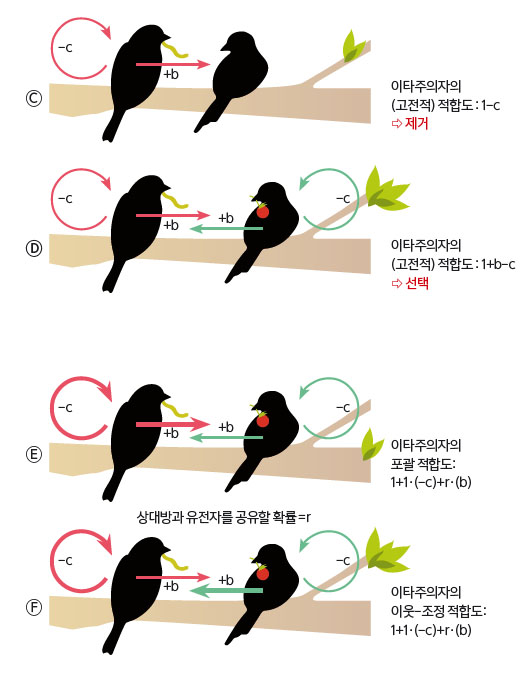
해밀턴은 사회적 상호작용을 올바르게 기재하는 방법을 찾았다. 그것도 두 개나 말이다(그림 3). 한 가지 방법은 각 개체를 ‘행위자’로 간주하는 것이다(그로부터 영향을 받는 수용자는 개체군 내의 다른 개체들과 자기 자신이 된다). 또 다른 방법은 각 개체를 ‘수용자’로 간주하는 것이다(그에게 영향을 주는 행위자는 개체군 내의 다른 개체들과 자기 자신이 된다).
사회적 행동을 보는 두 가지 방법
첫 번째 방법이 바로 ‘포괄 적합도(inclusive fitness)’다. 사회적 행동의 진화를 설명할 때, 남이 낳은 자식들 가운데 일부는 나 때문이고 내가 낳은 자식들 가운데 일부는 남 때문이라 골치 아프다고 하지 않았는가? 그러니 어떤 특정한 개체가 (자기 자신을 포함해) ‘다른 개체들에게 준’ 적합도상의 효과들만 오롯이 따지자는 것이다. 행위자와 수용자가 사회적 행동을 일으키는 유전자를 공유할 확률, 곧 ‘유전적 근연도(genetic relatedness)’로 각각의 적합도 효과를 일일이 가중한 다음에 모두 더한 값이 포괄 적합도다. 이 때, 남들이 나에게 준 적합도상의 효과들은 깡그리 무시한다.
이타주의자가 한 명과 상호작용하는 경우의 포괄 적합도를 계산해 보자(그림 Ⓔ). 수용자는 몇 명일까. “방금 한 명과 상호작용 한다며!”라고 투덜대며 한 명이라고 답했다면, 틀렸다. 수용자는 두 명이다. 행위자 자신과 상대방, 이렇게 둘이다. 행위자가 자기 자신과 유전자를 공유할 확률은 1이다. 이타적 행동으로 c만큼 적합도가 하락하므로, 1·(-c)가 이타적 행동이 행위자의 직접 적합도에 끼친 영향이다.
한편, 행위자가 상대방과 유전자를 공유할 확률을 r이라 하자. 이타적 행동으로 b만큼 상대방의 적합도가 상승하므로 r·(b)가 이타적 행동이 행위자의 간접 적합도에 끼친 영향이다. 기본으로 받는 적합도 1을 더하면 1+ 1·(-c) + r·(b)가 이타주의자의 포괄 적합도다(포괄 적합도는 직접 적합도와 간접 적합도의 합이라고 지난 시간에 이야기했다). 포괄 적합도가 1보다 크면, 이 이타적 행동은 자연 선택된다. 즉, 유전자의 관점에서는 –c+rb >;0이면 이타적 행동을 통해서 다음 세대에 남기는 복제본의 개수를 증가시킬 수 있다. 이를 ‘해밀턴의 규칙’이라 한다.
두 번째 방법은 ‘이웃-조정 적합도(neighbor-modulated fitness)’다. 이번에는 자기 자신을 포함해 다른 개체들이 ‘자신에게 준’ 적합도상의 효과들만 따진다. 각각에 맞는 유전적 근연도로 각각의 적합도 효과를 가중한 다음에 모두 더한 값이 이웃-조정 적합도다. 이 때, 내가 남들에게 준 적합도상의 효과들은 일체 무시한다.
이타주의자가 한 명과 상호작용하는 경우의 이웃-조정 적합도를 계산해 보자(그림 Ⓕ). 수용자는 이타주의자 한 명이다. 행위자는 이타주의자 자신과 상대방, 이렇게 두 명이다. 이타주의자가 자기 자신과 유전자를 공유할 확률은 1이다. 나 자신(행위자)의 이타적 행동으로 c만큼 나(수용자)의 적합도가 하락한다. 상대방(행위자)이 베푼 이타적 행동으로 b만큼 나의 적합도가 상승한다. 이 값에 근연도 r을 곱해서 가중하면 r·(b)가 상대방의 이타적 행동이 나의 적합도에 실제로 끼친 영향이다. 기본으로 받는 적합도 1을 더하면 1+ 1·(-c) + r·(b)가 이타주의자의 이웃-조정 적합도다. 이 값이 1보다 크면, 이 이타적 행동은 자연 선택된다. 즉, 포괄 적합도의 경우와 마찬가지로, –c+rb >;0이면 이타적 행동을 일으키는 유전자는 선택될 수 있다.
요약하자면, 포괄 적합도와 이웃-조정 적합도는 같은 사회적 상호작용을 보는 두 가지 동등한 방법이다. 둘 다 해밀턴의 규칙이라는 같은 결과를 낸다. 둘 다 사회적 상호작용을 만드는 유전자가 남기는 복제본의 개수를 정확히 세도록 해준다. 즉, 자연 선택은 각 개체의 포괄-적합도(혹은 이웃-조정 적합도)를 최대화한다.
여기까지 이해했다면 일단 핵심은 파악한 셈이다. 다음 화에서 해밀턴의 법칙과 유전적 근연도를 좀 더 탐구해 보자.