갈라파고스 중앙해령에서 처음 발견한 심해 열수생태계는 놀라움과 함께 지적 호기심을 자극했다. 둥그런 현무암 틈에 빼곡히 들어차 20~30cm까지 자란 대형 조개와 홍합, 키가 2m에 이르는 대형 관벌레, 이 고착동물 표면에 가득 붙어 사는 손톱보다 작은 삿갓조개, 이곳저곳을 기어 다니며 먹이를 찾는 흰 게, 세균들이 바위 표면에 대량으로 성장하여 카펫 같은 모습….
이 심해 생물들의 정체는 무엇일까? 이들은 어떻게 번성했을까? 빛이 들어오지 않는 캄캄한 심해에서 어떻게 영양을 섭취할까? 또 열수가 뿜어내는 유독한 황화수소(H2S)와 고농도 중금속의 극한 환경에서 생존할 수 있을까? 이 생물들은 다른 바다 밑에서도 살고 있을까? 이 생물들은 어디에서 기원하였을까? 해답을 찾기 위해 과학자들은 전 세계 바다 밑 다른 지역들로 심해탐사를 확장시켜나갔다. 이 같은 노력을 통해 열수생태계라는 큰 퍼즐이 한 조각씩 맞춰지고 있다.
태양 없이도 생존하는 화학합성생태계
심해에서 고압을 받은 바닷물은 해저 지각의 갈라진 틈을 따라 지하로 침투한다. 이 바닷물은 마그마의 열을 받아 뜨겁게 데워진 뒤, 지각의 갈라진 틈을 따라 다시 배출되는 순환 과정을 거친다. 이 과정에서 해양 지각에 함유된 다양한 광물과 화합물들이 열수에 혼합되거나 녹아 함께 상승한다. 육상의 온천과도 비슷해 보이는 심해 열수지역은 열수가 나오는 굴뚝처럼 생긴 분화구와 이들 주변으로 비교적 따듯한 환경(10~20℃)이 조성된 현무암 혹은 퇴적층 지대로 구성되어 있다. 열수분화구에서 바로 분출되어 나오는 열수 온도는 지역에 따라 차이가 있지만 보통 약 300℃다(2006년 남대서양 적도부근 한 열수구 입구에서 측정된 407℃가 현재까지 최고 기록).
굴뚝 모양의 열수분화구는 지각 틈 사이로 솟구친 열수가 찬 바닷물과 섞이는 순간 침전이 생기면서 형성된다. 마치 검은 연기를 뿜어 내는 것처럼 보여 ‘블랙스모커’라고 부른다. 그러나 열수가 모두 검은색을 띠는 것은 아니다. 열수는 분화구 외에도 지각 틈으로 느리게 새어나와 확산되기도 한다. 이 과정에서 열수에 포함된 많은 이온과 화합물들도 퍼져나간다. 바로 이 열수에 포함된 가스와 화합물들이 열수생태계를 지탱하는 에너지 원천이다.
열수 지역에서 화학합성은 생명 현상과 밀접한 관련이 있다. 독립영양세균이 황화수소나 메탄(CH4) 같은 환원성 화합물을 산화시켜 에너지를 획득하는 과정과 이산화탄소(CO2)나 메탄에 있는 무기 탄소를 유기물로 고정하는 대사 과정이다. 두 가지의 생명 현상은 세균과 열수에 사는 무척추동물 사이를 공생 관계로 엮어준다.
예를 들어 열수장님새우의 등껍질에는 화학합성공생세균이 가득히 붙어 사는데 장님새우는 이 공생세균을 먹이원으로 삼는다. 물론 열수 가까이서 헤엄치는 장님새우 덕분에 새우 몸에 붙어사는 세균은 생존에 필요한 화합물을 얻을 수 있다.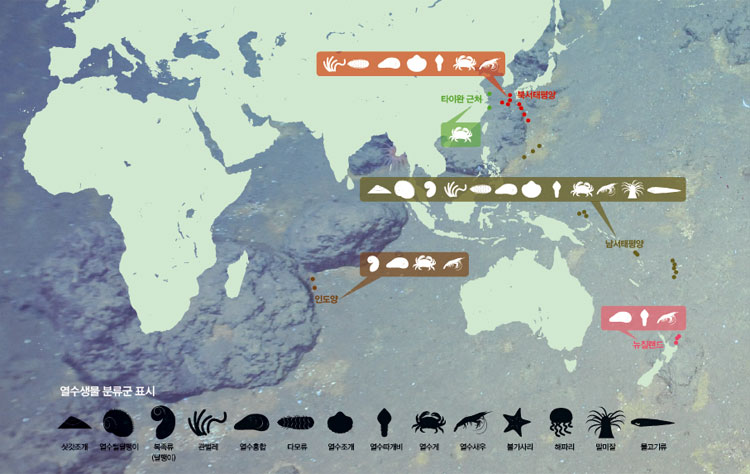
먹지 않아도 살 수 있는 열수생물
공생세균은 숙주 동물의 특별한 세포 안에 존재하기도 하는데 이러한 관계를 세포내공생이라 부른다. 열수관벌레는 관의 거의 대부분을 차지하고 있는 트로포솜이라는 기관에, 조개나 홍합 같은 이매패류와 해양달팽이 종류인 복족류에서는 아가미 조직의 박테리오사이트 세포 안에 공생세균이 산다. 이들은 물질대사를 통해 합성한 유기물을 숙주에게 영양분으로 준다.
이 같은 영양방식은 태양에너지를 이용해 광합성을 하여 양분을 얻는 식물이나 식물성 플랑크톤에 기반을 둔 해양 표층생태계와는 전혀 다르다. 화학합성 독립영양생물인 세균의 영양 공급은 숙주가 스스로 먹이를 섭취할 필요성을 줄이고, 극단적으로는 전혀 먹지 않아도 살 수 있게 만드는 등 다양한 적응을 유도했다. 예를 들어 열수관벌레 종은 소화기관이 아예 없다. 트로포솜을 가득 채운 세균이 주는 영양분만으로도 충분히 살아갈 수 있기 때문이다. 이매패류인 홍합은 유기물이나 미생물을 걸러 먹어 얻은 영양분과 공생세균이 제공하는 영양분을 흡수한다.
화학합성생태계는 전혀 다른 환경에서도 나타난다. 1983년 멕시코 만 플로리다 근처 급경사면의 심해저에서 새로운 종류의 심해 생태계가 앨빈 호 탐사로 우연히 발견됐다. 이 생태계는 주변 해수처럼 수℃ 수준의 차가운 온도 환경을 가져 냉수생태계라고 불린다. 이 곳은 해저에 쌓인 침전층에서 황화수소나 메탄, 탄화수소가 풍부한 유체가 스며 나온다. 덕분에 열수지역 생태계처럼 화학합성에 기반을 둔 관벌레나 홍합 같은 생물이 살아가고 있다.
화학합성생태계의 또 다른 유형은 해저에 가라앉은 고래 사체에서도 발견됐다. 앨빈 호는 1987년 캘리포니아 앞 바다 심해에서 고래 사체를 발견했는데, 고래 사체의 유기물이 미생물에 의해 분해되면서 환원성 화합물이 많은 환경이 조성됐고 이 환경에 적응한 생물종이 군집을 이룬 것이다.
이처럼 주변 환경에 비해 환원성 화합물의 농도가 높고, 이 환경에 특화된 세균의 화학합성이 존재할 경우 새로운 생태계가 만들어질 수 있다. 게다가 서로 다른 이유로 만들어진 화학합성생태계라도 그곳에 사는 동물들은 서로 진화 연관성이 높다고 추정할 수 있다. 서로 다른 서식지 별 생물종이 과거 어느 시점에선가 어느 한 공동조상에서 유래되어 오늘날 각각 다른 환경에 적응 진화해 살고 있다는 의미다.

열수생태계의 수명은 100~1만 년
최초의 열수생태계 발견에서 오늘날까지 약 35년이 경과하는 동안 과학자들의 탐사는 전 세계 바다로 확장되어 갔다. 이 생태계는 전세계 바다 밑 해양지각판과 판들이 확장하고 있는 중앙해령, 판들이 수렴하고 있는 섭입대의 호상분지, 그리고 열수작용이 있는 해산 등에 존재한다. 따라서 지리적으로 열수생태계는 마치 고립된 섬들처럼 여기저기 흩어져 분포한다. 한 열수구와 이웃하는 열수구가 수백에서 수천 km 간격을 두는 경우도 있고, 중앙해령의 축이 변환단층으로 끊기면서 빗겨나는 경우도 많다.
이 때문에 지금까지 발견된 열수생물들은 매우 높은 지역고유성을 보인다. 평균적으로 열수생물종의 약 70%가 특정 지역에서만 발견되고 다른 지역에서는 발견되지 않는다. 냉수생태계 종들도 지역고유성이 40%나 된다. 열수서식지의 고립과 파편화에 따른 자연스러운 귀결이다. 지리적으로 격리되어서 종분화가 일어난 것이다.
한 지역의 열수 활동이 영원하지 않다는 점도 열수생물들의 지역고유성을 만든다. 열수 서식지는 시간적으로 보면 해저 마그마 활동에 영향을 받으면서 생성과 소멸의 주기를 갖는다. 열수에 포함된 황화수소나 메탄의 농도는 생성된 지 얼마 안 된 뜨거운 열수구에서 가장 높으며 시간이 지날수록 점점 감소하다 너무 낮아지면 열수생태계는 결국 사멸하게 된다. 열수작용의 존재 기간은 매년 18cm 이상 움직이는 남동태평양 중앙해령 같은 곳에서는 약 100년, 연당 0.6cm 정도 움직이는 북대서양 중앙해령에서는 약 1만 년을 넘을 정도로 차이가 크다. 이처럼 시공간적 변화가 큰 환경에 맞서서 열수생물은 높은 번식력과 분산능력 그리고 빠른 개체성장 속도로 적응 진화했다. 이런 능력을 갖추지 못했다면 어느 시점 어느 지역에선가 멸종하고 말았을 것이다.
1991년 4월 동태평양 중앙해령 북위 9˚상의 열수구를 탐사하던 미국 럿거스대 열수생물 연구진은 일어난 지 얼마 안된 해저 화산폭발과 용암 분출이 초래한 광경을 앨빈 호에서 직접 목격했다. 자연이 준 천우일회의 기회였다. 이전에 존재하던 열수생물들은 화산폭발로 용암과 퇴적물 더미에 묻혀버렸고 새로 뿜어져 나오는 열수 주변에는 세균이 대량으로 증식해 만들어진 흰 눈 같은 덩어리가 바람에 날리듯이 주변 해수층에 가득했다.
이곳은 열수생태계의 천이가 어떻게 이루어지는지를 알려주는 장기 모니터링 연구지가 되었다. 열수서식지의 천이 과정은 매우 빨랐다. 초기 화산폭발 이후 세균 매트가 번성하고 이 세균 매트는 새우, 게, 물고기 등 다른 청소부들을 유인했다. 1년 뒤 세균 매트는 줄어들고 작은 관벌레인 테브니아(Tevnia jerichonana)가 정착했으며, 1년 8개월 뒤에는 대형 관벌레(Riftia pachyptila)가 테브니아보다 훨씬 많이 성장했다. 2년 6개월 후 대형 관벌레 한편으로 새로운 개체들도 계속 정착해 자랐다. 이 기간에 홍합들이 나타나기 시작했으며 종의 수가 늘어났다. 3년 7개월 후 대형 관벌레가 2000개체 이상이 되었으며 그 전 해에 정착했던 홍합들이 서식지를 우점했다.


우리나라가 발견한 남극 열수 생태계
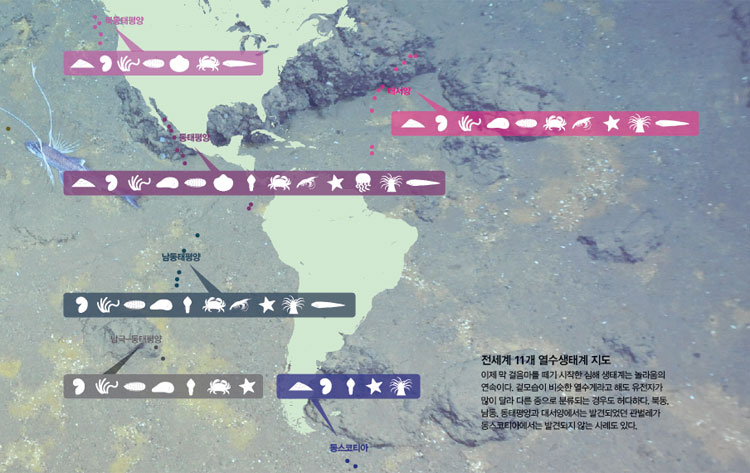
오늘날 전 세계 바다 밑에서 발견된 열수구는 약 200군데에 이른다. 이 가운데는 유인잠수정이나 원격조종 잠수정으로 직접 생물 시료를 채집한 지역도 있고 심해 카메라나 화학센서로 열수의 작용만 확인한 곳도 있다. 열수 탐사 현황을 볼 수 있는 인터넷 사이트는 중앙해령 연구를 위한 국제협력기구(www.interridge.org)가 있다. 우리나라 연구진(대표 극지연구소 박숭현 박사)도 회원국으로 참여하고 있다. 특히 2011년까지 관측된 열수지점을 정리해둔 파일이 제공돼 누구나 구글 지도를 통해 열수 지점을 볼 수 있다.
열수생물 연구는 35년밖에 안 되는 상대적으로 짧은 역사라서 지도를 살펴보면 광대한 해역이 아직도 연구자의 손길이 닿지 않은 채로 남겨져 있다. 지구의 중앙해령 시스템은 북동태평양, 동태평양, 서태평양, 남서태평양, 서태평양, 인도양과 남북극에 걸쳐 그 길이가 총 6만km에 달한다.
지금까지 생물조사가 이루어진 지역의 열수생물을 비교해보면 지구 전체의 열수생태계는 11개의 생물구역으로 나눠볼 수 있다. 그러나 이 숫자는 미래에 추가될 열수구 발견에 의해 변할 수 있다. 수치로 보면 중앙해령 6만km 구간 중 탐사가 이루어진 구간은 약 20%밖에 안 된다. 더구나 탐사 지점 간에는 약 100 km의 간격(조사 미구간)이 존재한다. 그러므로 새로운 열수구와 그곳에 서식하는 새로운 종들 그리고 이미 밝혀진 열수생물의 자매 집단들이 미래에도 계속 발견될 것이다. 잃어버린 퍼즐조각들이 하나하나 찾아지듯이.
우리나라는 2011년 태평양과 남극 사이에 위치하는, 지구 중앙해령 시스템에서 제일 긴 구간이면서도 지금까지 연구가 가장 안된, 그래서 세계 과학계의 초미의 관심 대상인 태평양-남극 중앙해령 탐사에 도전했다. 고위도 남극 바다의 험난한 파도와 날씨를 무릅쓰고 이때부터 2013년까지의 후속 탐사를 통해 세계 최초로 남극의 가장 고위도 지점(남위 62˚)에서 열수구와 열수생물의 과학적 증거를 찾아냈다. 자세한 발견 내용은 곧 논문으로 발표될 예정이다. 쾌거가 아닐 수 없다.