대부분의 곤충은 그들의 활동시간대를 1일중 어느 특정시간대에 두고 있다. 이는 곤충의 어디엔가 시계로서의 기능을 가진 구조가 존재하고 있다는 말이 된다.
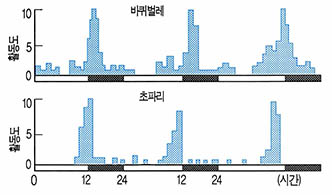
(그림1)은 야행성 곤충인 바퀴벌레와 주행성 곤충인 초파리의 활동리듬을 나타낸 것이다. 바퀴벌레는 하루 중 일몰 직후 몇시간 이내에 최대의 활동치를 나타내고 그 이후 서서히 활동이 줄어들었다. 반면 초파리는 일출직전에 최대치의 활동을 나타낸 후 암기에 정지하고 있다. 이러한 활동리듬은 실험실의 인위적 명-암 환경(12L:12D)의 외인성 조건 하에서 얻어진 결과이다.
곤충에 있어서 광주성(photoperiodism)은 개일 중심구동체(驅動禮:central circadian pacemake), 즉 주시계(主時計: master clock)라는 개념에 의하여 설명되고 있다. 이 개념은 이를 긍정할 수 있는 많은 실험결과에 의해 타당성이 인정되고 있다.
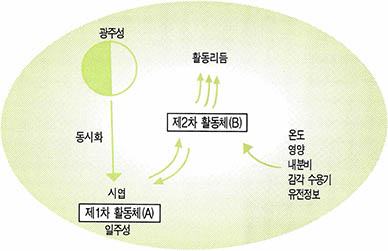
여기에는 개일 구동제와 같은 커다란 집합체와 그로부터 작동한 보다 다양한 리듬이 존재하며 이들이 전체로서 하나의 개일 계(System)를 형성하고 있다. 바퀴벌레와 같은 경우 (그림2)에서와 같이 개일 계가 제 1활동체(primary oscillator)와 제 2활동체(secondary oscillator)로 구분하고 있다.
즉 시엽(視葉)은 24시간의 외인성 광주기를 제 1활동체에서 동조시켜 그 정보를 제 2활동체로 넘겨준다. 제 2활동체는 그 정보대로 활동하는 것이 아니고 온도 영양 내분비 감각기능 유전정보 등의 조절기능과 일치시켜 그 결과로 활동을 하게 된다는 것이다.
바퀴벌레의 활동리듬
바퀴벌레를 24시간 주기에서 명암주기 12:12로 조절해두면 조명이 꺼진 직후부터 행동을 개시하여 몇시간동안 활발히 행동한 후 서서히 활동이 낮아지고, 이어서 조명이 켜지면 행동은 극도로 낮아지고 결국 정지한다.
이러한 활동리듬은 정확하게 24시간의 광주기에 동조하여 반복된다. 그러나 이들을 연속DD(암흑) 또는 연속LL(조명)의 조건에 두면 매일의 주기성을 나타내지만 개체고유의 시간에 의한 리듬이 나타난다.
우리들 주변에서 흔히 볼 수 있는 바퀴벌레의 48일간에 걸친 활동리듬을 조사해보았다. 제 1일째부터 제 27일째까지 연속 암환경 하에서의 '자유연속리듬을 나타내 본 결과 바퀴벌레는 매일이 23시간30분으로 하루 30분이 짧은 주기를 보여주고 있다. 한편 제 28일째부터 매일 명-암 12:12의 조건으로 둔 결과 정확하게 암기가 시작되는 시각에 맞추어 24시간 주기의 리듬이 형성되었다.
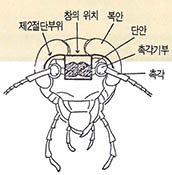
이와 같이 광주기가 활동리듬을 동조시키는 역할을 한다면, 이러한 빛의 정보는 바퀴벌레 몸의 어떤 부분인가를 통하여 내부의 시계로 전달되는 것으로 추정할 수 있다. 곤충의 두부에는 복안, 단안, 촉각, 뇌, 뇌간, 알라타체 등의 주요기관들이 있다.
바퀴벌레를 C${O}_{2}$로 마취한 후 두부를 고정하여 해부 현미경 밑에서 외과적 수술을 행하였다. 수술은 (그림5)의 4각형 부분을 예리한 면도날을 사용하여 피하조직을 손상하지 않게 절단하여 적출했다. 그러면 (그림6)에서와 같이 전대뇌(Br.I)와 중대뇌(Br.Ⅱ)의 대부분이 노출된다.
원하는 각 부위의 수술을 행한 후 보존해 둔 4각형의 피부를 원래의 부위에 부착시킨 후 파라핀으로 봉하여 활동리듬이 생성되는지 여부를 살펴보았다.
외과적 수술을 행한 결과 1)양측의 단안을 모두 적출하여도 보행활동의 리듬은 존속하였다. 2)복안의 개안과 시엽을 연결하는 시신경을 절단한 경우 명-암 12:12주기에 동조하지 않고 자유연속리듬이 존속되었다. 3)양측의 시신경을 적출하고 수술 부위에 피부 대신 투명막을 설치하여 빛이 직접 뇌간부에 도달하도록 해도 명-암 12:12주기에 동조하지 않고 자유연속리듬이 나타났다.
이러한 결과를 통해 외부 빛 정보는 개안으로부터 시신경을 거쳐 뇌로 전해져서 행동의 리듬을 조정하는 것으로 판단하였다. 복안이 외부 빛의 정보를 제공한다면 생체시계 그 자체의 존재가 어디인가를 추적하지 않을 수 없다.
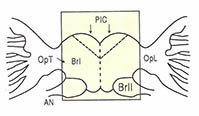
곤충의 뇌에서 시신경과 직접 연결되는 부위로 뇌간부(PIC)가 있다. (그림6)의 뇌간부(그림의 사각형 부분)의 각 부분을 절제하여 분석한 결과에서도 부분적인 자유연속리듬이 존속하였다. 그러나 시신경간-전대뇌의 주엽과 시엽과를 연결하는 교(橋)의 부분을 절제한 결과 자유연속리듬이 완전히 회실되었다. 즉 시엽이 생체시계의 구동(驅動)역할을 하는 기관임을 알 수 있었다.
위의 실험 외에 수많은 연구자들의 끈질긴 생체시계 추적실험이 이루어져왔다. 그 결과 1960년에 이르러 하커(Harker)에 의해 생체시계의 존재 부위가 식도하 신경구(suboesophageal ganglidon)에 있는 신경분비세포의 분비 사이클이 내인적 생체리듬을 규제하여 제 1단계의 시계를 구성하고, 이 시계는 다시 측심체(corpora cadiaca)로 연결되는 제 2의 시계로 전달된다고 하였다.
한편 로버트(Robert, 1970)는 뇌간부에 다수 존재하는 신경분비세포가 시계의 구동역할을 한다고 주장하기도 하였다.
바퀴벌레의 생체시계 구조
프린스턴 대학의 피텐드리히 교수 연구실은 여러 형태의 실험을 행하여 바퀴벌레의 성충행동을 지배하는 체내시계의 구조를 밝혔다.
(그림3)은 이들이 밝힌 체내시계의 모식도로서, 시계는 전대뇌에 있으며, 시엽과 뇌간부의 두 부분으로 되어 있고, 여기에 좌우 양반구 이중으로 보정된 구조를 가지고 있다. 시엽이 시계의 구동 역할을 하는 자리이며, 여기서부터 나온 정보는 신경로를 따라 중개(仲介)의 자리인 뇌간부로 전해진다.
뇌간부에 있는 신경분비세포가 어떤 종류의 '활동억제 호르몬'을 시엽의 정보에 의하여 주기적으로 분비하고, 이것이 체액을 통하여 흉부에 있는 신경구의 활동수준과 운동의 주기성을 조절하는 것으로 결론지었다.
이들은 결론으로서 시엽은 외계의 조건과는 무관하게 약 24시간의 리듬을 가지고 활동하는 체내시계의 중추적 역할을 하는 기관임을 강조하고 있다. 이 시엽은 좌우 한쌍으로 양 시엽간을 연결하고 있는 신경연락이 있기 때문에 두개의 구동체는 동일한 위상으로 작동하게 되고, 따라서 양 구동체 간에 정보의 오차가 일어나지 않는다.
결과적으로 외계의 빛 시그널은 복안으로부터 시신경을 거쳐 시엽에 직접 전해지고, 여기에서 외계의 빛의 조건이 파악되어 구동체는 정확한 시간을 맞추게 되는 것이다.