개화조절 유전자에 대한 연구는 최근 실험모델인 애기장대 식물에서 빠른 속도로 진행돼 왔다. 연구결과 개화와 관련된 많은 유전자들이 밝혀졌는데, 이 가운데 중요한 기능을 하는 몇몇 유전자만 간단히 살펴보자.
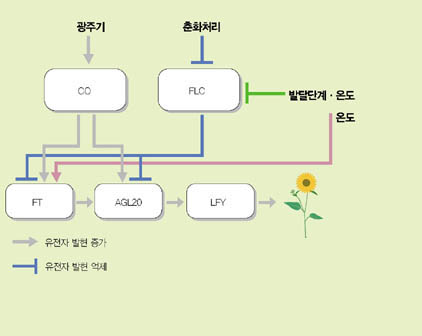
LEAFY(LFY)라는 유전자는 개화조절 유전자 명령체계 중 가장 하위에서 작용하는 유전자로 꽃의 발생을 직접 조절하는 기능을 가진다. 그 위에 FT와 AGL20이라는 두개의 유전자가 있고, 이들 유전자의 발현을 조절하는 상위 유전자 CONSTANS (CO)와 FLC가 있다. 흥미롭게도 CO와 FLC는 서로 반대되는 역할을 한다. 즉 CO는 FT와 AGL20 유전자의 발현을 증가시키는 기능을 하는 반면, FLC는 각각의 유전자 발현을 억제시키는 기능을 한다. FLC는 사실상 개화억제자인 셈이다. FLC를 제외한 나머지 유전자들은 그 발현량이 증가하면 개화가 촉진되고, 발현량이 감소하면 개화가 지연된다. 반면 FLC의 경우에는 발현량이 증가하면 개화가 지연되고, 발현량이 감소하면 개화가 촉진된다.
이같은 결과는 국내외 관련학자들의 연구성과다. ‘네이처 지네틱스’ 2월호에 개화유전자와 관련된 논문을 발표했던 고려대 안지훈 박사팀은 FT를, 필자가 이끄는 연구팀은 AGL20을 연구했다. 영국 존인네스센터의 커플랜드 박사팀은 CO를, 미국 스크립스연구소의 케이 박사팀은 CO와 생물시계, 그리고 여러 유전자와의 관계를, 미국 위스콘신대의 아마시노 교수팀은 FLC를 각각 연구했다.
개화조절 유전자는 장일식물인 애기장대뿐만 아니라 단일식물인 벼에서도 나타나고 있고, 그 기능 또한 유사한 것으로 현재 보고되고 있다. 따라서 개화조절 유전자 네트워크는 거의 모든 식물에 적용될 것으로 보인다.
식물이 제철인지 아는 방법
이제 계절에 따라 식물이 어떻게 개화시기를 조절하는지 알아보자. 계절에 따라 광주기와 온도가 규칙적으로 변화하고, 식물은 이들 변화를 인지해 제철에 꽃을 피운다.
먼저 광주기에 따라 개화시기가 조절되는 이유는 무엇일까. 간단히 말하면 개화유전자 CO와 FT의 발현이 생물시계와 피토크롬에 의해 조절되기 때문이다.
FT 유전자는 그 발현량이 증가되면 AGL20과 LFY 유전자의 발현을 증가시켜 개화를 일으킨다. FT 유전자의 발현은 CO 유전자에 의해 조절되고, CO 유전자의 발현은 다시 광주기를 인지하는 생물시계의 조절을 받고 있다.
장일식물인 애기장대에서 밝혀진 개화조절 모델을 구체적으로 살펴보자(그림5).
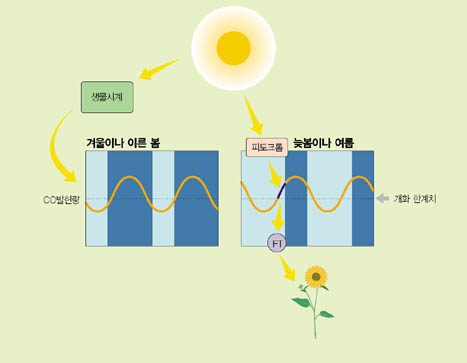
CO 유전자의 발현량을 시간별로 측정해보면 24시간의 주기로 늘었다, 줄었다를 반복하고 있다.
그런데 흥미롭게도 계절에 따라 이 주기가 약간 어긋나는 현상을 볼 수 있다. 겨울이나 초봄에는 낮 동안 CO 유전자 발현량이 개화에 필요한 한계치(역치) 이하로 낮은 반면, 늦봄이나 여름에는 낮 동안 CO 유전자의 발현량이 역치 이상으로 올라가는 것을 볼 수 있다. 낮 동안의 이런 CO 유전자 발현량의 차이가 겨울이나 초봄에는 개화가 지연되고 늦봄이나 여름에는 개화가 촉진되는 원인이 된다. 다시 말하면 겨울이나 초봄의 경우 낮에는 CO의 양이 너무 적어서 FT 발현이 증가되지 못하고, 밤에는 CO의 양은 많지만 빛이 없어서 FT 발현이 증가되지 못하는 것이다. 반면 늦봄이나 여름의 경우 낮에 충분한 양의 CO가 있어서 피토크롬과의 상호작용에 의해 FT 발현을 증가시킬 수 있기 때문에 개화가 빨리 진행되는 것이다.
이처럼 애기장대와 같은 장일식물이 어떻게 계절을 인식하는지 명쾌하게 설명된다. 그러나 단일식물의 경우에도 이 모델이 적용될지는 좀더 두고 봐야 하겠다. 현재 단일식물의 실험모델로서 벼가 집중 연구되고 있으니 조만간 그 해답을 얻을 수 있을 것으로 기대된다.

개화억제 유전자 활동 막아야
다음으로 온도에 따른 개화시기의 조절은 어떻게 이뤄질까. 다시 개화조절 유전자 네트워크를 살펴보자.
춘화처리는 FLC라는 개화억제 유전자의 발현을 완전히 억제함으로써 개화를 유도하는 방법이다. 즉 FT와 AGL20 유전자의 발현을 억제하던 FLC 유전자의 역할이 춘화처리에 의해 사라짐으로써 FT와 AGL20 유전자의 발현량이 증가하고 결과적으로 개화를 유도할 수 있게 되는 것이다. 한편 주변의 적당히 낮은 온도를 인식하는 메커니즘은 FLC와 FT 유전자의 발현을 적절히 조절하는 것으로 알려져 있다.
따라서광주기, 온도, 겨울철 저온 등의 모든 환경요인이 결국 FT, AGL20, LFY 유전자의 발현을 조절함으로써 개화시기를 조절하는 것이다. 이것이 계절에 따라 식물이개화시기를 조절하는 방법이라 할 수 있다.